(Pilar valla chasco no se ven ni imagenes ni videos ni na, te lo llevaé en el USB a clase para verlo mejor ) v.v
miércoles, 24 de febrero de 2010
Ribosoma y cloroplastos
Ribosoma : Estructura, composición y demás
Los ribosomas fueron descritos por primera vez por Palade en 1953 al microscopio electrónico. Los estudió en
el R.E. rugoso por la cara no citosómica, y también libres.
En su composición entra el ARNr y el proteínas.
El ARNr (el 60%).Quedaría por fuera rodeando a las proteínas.
Están presentes en todas las células, situados en el citosol.. El núcleo carece de ellos. Se encuentran también en
mitocondrias y en plastos
.Los ribosomas de procariotas y eucariotas son muy similares. Cada ribosomas está formado por dos
subunidades, una pequeña (1) y otra grande(2) que se mantienen juntas formando un complejo.
La presencia de Mg2+ hace que ambas subunidades se unan. Su ausencia hace que se despeguen.
(1) (2)
Durante la síntesis de proteínas, la subunidad menor se une al ARNm y al ARNt. Mientras, la grande se
pega a los enlaces peptídicos.
Los componentes ribosomales se designan por los valores de S .
Un ribosoma procariota es de 70 S, siendo 50 S la subunidad mayor y de 20 S la menor .La subunidad grande presenta 3 salientes y está formada por una moléscula de ARN grande de 23S y una pequeña de ARN de 5 S. Además contiene 34 proteínas. La subunidad pequeña muestra 2 lóbulos y
muestra una molécula grande de ARN de 16 S, tiene 21 proteínas (todas distintas). Casi todas las proteínas
son ricas en aá. básicos.
Un ribosomaeucariota es de 80 S. Es un poco más grande que el de los procariotas. La subunidad mayor es de
55 S y la menor es de 25 S.
A pesar de las diferencias, ambos ribosomas tienen una estructura casi idéntica y una función muy parecida.
Los ribosomas tienen un alto número de proteínas. Se cree que las
proteínas ribosomales aumentan la función de los ARNr y también se cree que son las moléculas del ARNr las
que catalizan la mayoría de las reacciones del ribosoma y no las proteínas
El ADN ribosómico constituye el 70 % de todo el citoplasma.
Función principal:Síntesis de proteinas
Para la síntesis de proteínas se requiere la unión de una subunidad mayor y otra menor, pero no es necesario
que sean siempre las mismas. Al terminar de fabricar las proteínas, se separan. Cuando vuelven a sintetizar se
unen dos diferentes.
Para la síntesis de proteínas los ribosomas se asocian en grupos mediante un filamento de unos 2 nm. de
ARN−m, . Estos ribosomas asociados se denominan polisomas, y
suelen adoptar una figura en espiral. La subunidad menor queda hacia el interior de la espiral.
Los ribosomas forman polisomas para realizar cualquier tipo de síntesis proteicas: Para la síntesis de proteínas, los ribosomas recorren el ARNm desde un extremo a otro. Por cada tres nucleótidos recorridos, incorporan un aminoácido a la cadena de proteínas que están sintetizando;
aminoácidos que les proporciona el ARNt . Cuando han completado el recorrido, los ribosomas se liberan del
ARNm y suelta la proteína ya terminada. Mientras se esté sintetizando proteínas, por cada ribosoma que
abandona el polisoma en el extremo final, otro se incorpora en el inicial, de modo que el ritmo de trabajo del
polisoma siempre es complejo.
El ARNt y la cadena de aminoácidos que se está formando se encuentran en un canal situado en la subunidad
mayor.
Hay agentes que inhiben la síntesis de proteínas ribosomales que actúan en eucariotas y en procariotas. Por
ejemplo, la PUROMICINA y la ACTINOMICINA D.
El proceso se repite tantas veces como aás. se incorporen. Cada ARNt entrante lo hace en el lugar A.
se une el polipéptido en formación y al desplazarse 3 nucleótidos el ribosoma, se produce la liberación del
ARNt unido al polipéptido de formación y el último ARNt incorporado se desplaza de A al P para dejar
sitio al siguiente ARNt.
Aquí se muestra un video del proceso de dicha síntesis:
http://www.youtube.com/watch?v=FNqmh4PoMPQ
El final de la síntesis tiene lugar cuando se interrumpe la producción.
Una proteína denominada FACTOR DE LIBERACIÓN es la responsable de ello .
El factor de liberación hace que la peptidil tranferasahidrolice el peptidil ARNt en lugar de unirlo a otro aá., con lo que queda libre la proteína. que no puede unirse a él ningún otro aá
.
---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
Cloroplastos: Estructura y función
Estructura Interior de un cloroplasto,
Las dos membranas del cloroplasto poseen una diversa estructura continua que delimita completamente el cloroplasto. Ambas se separan por un espacio intermembranoso llamado a veces indebidamente espacio periplastidial. La membrana externa es muy permeable gracias a la presencia de porinas. Sin embargo no tanto como la membrana interna, que contiene proteínas específicas para el transporte.
La cavidad interna llamada estroma, en la que se llevan a cabo reacciones de fijación de CO2, contiene ADN circular, ribosomas , gránulos de almidón, lípidos y otras sustancias. También, hay una serie de sáculos delimitados por una membrana llamados tilacoides los cuales se organizan en los cloroplastos de las plantas terrestres en apilamientos llamados grana. Las membranas de los tilacoides contienen sustancias como los pigmentos fotosintéticos (clorofila, carotenoides, xantófilas) y distintos lípidos; proteínas de la cadena de transporte de electrones fotosintética y enzimas, como la ATP-sintetasa.
el cloroplasto tiene tres sistemas de mebrana, que forman tres compartimentos, el espacio intermembrana, el estroma y el espacio intratilacoidal
Funciones
Es el orgánulo donde se realiza la fotosíntesis. Existen dos fases, que se desarrollan en compartimentos distintos:
* Fase luminosa: Se realiza en la membrana de los tilacoides, donde se halla la cadena de transporte de electrones y la ATP-sintetasa responsables de la conversión de la energía lumínica en energía química (ATP) y de la generación poder reductor (NADPH).
* Fase oscura: Se produce en el estroma, donde se halla el enzima RuBisCO, responsable de la fijación del CO2 mediante el ciclo de Calvin.
Aquí se dan dos videos : uno alude al proceso de la fotosíntesis y el otro a una vista microscópica de estos cloroplastos
http://www.youtube.com/watch?v=BYWJHwF_yok(procesos fotosintesis)
http://www.youtube.com/watch?v=cjliUg8kicM&feature=related (microscopio)
CURIOSIDAD!
Se cree que los cloroplastos se han originado a partir de organismos procariotas fotosintéticos, de algas azules para ser mas exacto, que se convirtieron en células primitivas eucariotas aeróbicas por endosimbiosis. Esa simbiosis se cree que se dió hace más o menos 1,2 billones de años de ahí evolucionarían a algas rojas, después a algas pardas y verdes y a los vegetables superiores como actualmente sucede.
Los ribosomas fueron descritos por primera vez por Palade en 1953 al microscopio electrónico. Los estudió en
el R.E. rugoso por la cara no citosómica, y también libres.
En su composición entra el ARNr y el proteínas.
El ARNr (el 60%).Quedaría por fuera rodeando a las proteínas.
Están presentes en todas las células, situados en el citosol.. El núcleo carece de ellos. Se encuentran también en
mitocondrias y en plastos
.Los ribosomas de procariotas y eucariotas son muy similares. Cada ribosomas está formado por dos
subunidades, una pequeña (1) y otra grande(2) que se mantienen juntas formando un complejo.
La presencia de Mg2+ hace que ambas subunidades se unan. Su ausencia hace que se despeguen.
(1) (2)
Durante la síntesis de proteínas, la subunidad menor se une al ARNm y al ARNt. Mientras, la grande se
pega a los enlaces peptídicos.
Los componentes ribosomales se designan por los valores de S .
Un ribosoma procariota es de 70 S, siendo 50 S la subunidad mayor y de 20 S la menor .La subunidad grande presenta 3 salientes y está formada por una moléscula de ARN grande de 23S y una pequeña de ARN de 5 S. Además contiene 34 proteínas. La subunidad pequeña muestra 2 lóbulos y
muestra una molécula grande de ARN de 16 S, tiene 21 proteínas (todas distintas). Casi todas las proteínas
son ricas en aá. básicos.
Un ribosomaeucariota es de 80 S. Es un poco más grande que el de los procariotas. La subunidad mayor es de
55 S y la menor es de 25 S.
A pesar de las diferencias, ambos ribosomas tienen una estructura casi idéntica y una función muy parecida.
Los ribosomas tienen un alto número de proteínas. Se cree que las
proteínas ribosomales aumentan la función de los ARNr y también se cree que son las moléculas del ARNr las
que catalizan la mayoría de las reacciones del ribosoma y no las proteínas
El ADN ribosómico constituye el 70 % de todo el citoplasma.
Función principal:Síntesis de proteinas
Para la síntesis de proteínas se requiere la unión de una subunidad mayor y otra menor, pero no es necesario
que sean siempre las mismas. Al terminar de fabricar las proteínas, se separan. Cuando vuelven a sintetizar se
unen dos diferentes.
Para la síntesis de proteínas los ribosomas se asocian en grupos mediante un filamento de unos 2 nm. de
ARN−m, . Estos ribosomas asociados se denominan polisomas, y
suelen adoptar una figura en espiral. La subunidad menor queda hacia el interior de la espiral.
Los ribosomas forman polisomas para realizar cualquier tipo de síntesis proteicas: Para la síntesis de proteínas, los ribosomas recorren el ARNm desde un extremo a otro. Por cada tres nucleótidos recorridos, incorporan un aminoácido a la cadena de proteínas que están sintetizando;
aminoácidos que les proporciona el ARNt . Cuando han completado el recorrido, los ribosomas se liberan del
ARNm y suelta la proteína ya terminada. Mientras se esté sintetizando proteínas, por cada ribosoma que
abandona el polisoma en el extremo final, otro se incorpora en el inicial, de modo que el ritmo de trabajo del
polisoma siempre es complejo.
El ARNt y la cadena de aminoácidos que se está formando se encuentran en un canal situado en la subunidad
mayor.
Hay agentes que inhiben la síntesis de proteínas ribosomales que actúan en eucariotas y en procariotas. Por
ejemplo, la PUROMICINA y la ACTINOMICINA D.
El proceso se repite tantas veces como aás. se incorporen. Cada ARNt entrante lo hace en el lugar A.
se une el polipéptido en formación y al desplazarse 3 nucleótidos el ribosoma, se produce la liberación del
ARNt unido al polipéptido de formación y el último ARNt incorporado se desplaza de A al P para dejar
sitio al siguiente ARNt.
Aquí se muestra un video del proceso de dicha síntesis:
http://www.youtube.com/watch?v=FNqmh4PoMPQ
El final de la síntesis tiene lugar cuando se interrumpe la producción.
Una proteína denominada FACTOR DE LIBERACIÓN es la responsable de ello .
El factor de liberación hace que la peptidil tranferasahidrolice el peptidil ARNt en lugar de unirlo a otro aá., con lo que queda libre la proteína. que no puede unirse a él ningún otro aá
.
---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
Cloroplastos: Estructura y función
Estructura Interior de un cloroplasto,
Las dos membranas del cloroplasto poseen una diversa estructura continua que delimita completamente el cloroplasto. Ambas se separan por un espacio intermembranoso llamado a veces indebidamente espacio periplastidial. La membrana externa es muy permeable gracias a la presencia de porinas. Sin embargo no tanto como la membrana interna, que contiene proteínas específicas para el transporte.
La cavidad interna llamada estroma, en la que se llevan a cabo reacciones de fijación de CO2, contiene ADN circular, ribosomas , gránulos de almidón, lípidos y otras sustancias. También, hay una serie de sáculos delimitados por una membrana llamados tilacoides los cuales se organizan en los cloroplastos de las plantas terrestres en apilamientos llamados grana. Las membranas de los tilacoides contienen sustancias como los pigmentos fotosintéticos (clorofila, carotenoides, xantófilas) y distintos lípidos; proteínas de la cadena de transporte de electrones fotosintética y enzimas, como la ATP-sintetasa.
el cloroplasto tiene tres sistemas de mebrana, que forman tres compartimentos, el espacio intermembrana, el estroma y el espacio intratilacoidal
Funciones
Es el orgánulo donde se realiza la fotosíntesis. Existen dos fases, que se desarrollan en compartimentos distintos:
* Fase luminosa: Se realiza en la membrana de los tilacoides, donde se halla la cadena de transporte de electrones y la ATP-sintetasa responsables de la conversión de la energía lumínica en energía química (ATP) y de la generación poder reductor (NADPH).
* Fase oscura: Se produce en el estroma, donde se halla el enzima RuBisCO, responsable de la fijación del CO2 mediante el ciclo de Calvin.
Aquí se dan dos videos : uno alude al proceso de la fotosíntesis y el otro a una vista microscópica de estos cloroplastos
http://www.youtube.com/watch?v=BYWJHwF_yok(procesos fotosintesis)
http://www.youtube.com/watch?v=cjliUg8kicM&feature=related (microscopio)
CURIOSIDAD!
Se cree que los cloroplastos se han originado a partir de organismos procariotas fotosintéticos, de algas azules para ser mas exacto, que se convirtieron en células primitivas eucariotas aeróbicas por endosimbiosis. Esa simbiosis se cree que se dió hace más o menos 1,2 billones de años de ahí evolucionarían a algas rojas, después a algas pardas y verdes y a los vegetables superiores como actualmente sucede.
Anatomia de las mitocondrias
Anatomia de las mitocondrias
Las mitocondrias son estructuras muy plásticas que se deforman, se dividen y fusionan. Normalmente se las representa en forma alargada. Su tamaño oscila entre 0,5 y 1 μm de diámetro y hasta 7 μ de longitud.Su número depende de las necesidades energéticas de la célula. Al conjunto de las mitocondrias de la célula se le denomina condrioma celular.
Las mitocondrias, orgánulos esféricos o alargados, aparecen repartidas por el citoplasma de las células eucarióticas . Una mitocondria está formada por una doble membrana de estructura similar a la de la membrana citoplasmática. La membrana mitocondrial externa es lisa, pero la interna se dispone formando una serie de pliegues llamadas crestas. El centro de la mitocondria se denomina matriz. Además poseen un espacio intermembranoso.Algunas enzimas implicadas en la respiración celular, incluyendo las que forman ATP, están localizadas sobre las crestas de la membrana interna mitocondrial y muchos de los pasos metabólicos relacionados con la respiración celular se realizan en la matriz.
Su número varía ampliamente según el tipo de organismo o tejido. A lgunas células carecen de mitocondrias o poseen sólo una, mientras que otras pueden contener varios miles.Este orgánulo se compone de compartimentos que llevan a cabo funciones especializadas. Las proteínas mitocondriales varían dependiendo del tejido y de las especies: en humanos se han identificado 615 tipos de proteínas distintas en mitocondrias de músculo cardíaco;mientras que en ratas se han publicado 940 proteínas codificadas por distintos genes. Se piensa que el proteoma mitocondrial está sujeto a regulación dinámica.Aunque la mayor parte del ADN de la célula está en el núcleo celular, la mitocondria tiene su propio genoma, que muestra muchas semejanzas con los genomas bacterianos.
lgunas células carecen de mitocondrias o poseen sólo una, mientras que otras pueden contener varios miles.Este orgánulo se compone de compartimentos que llevan a cabo funciones especializadas. Las proteínas mitocondriales varían dependiendo del tejido y de las especies: en humanos se han identificado 615 tipos de proteínas distintas en mitocondrias de músculo cardíaco;mientras que en ratas se han publicado 940 proteínas codificadas por distintos genes. Se piensa que el proteoma mitocondrial está sujeto a regulación dinámica.Aunque la mayor parte del ADN de la célula está en el núcleo celular, la mitocondria tiene su propio genoma, que muestra muchas semejanzas con los genomas bacterianos.
Las mitocondrias tienen las crestas laminares o digitiformes.
La mayoría son de crestas laminares, mientras que las mitocondrias de células productoras de estrógenos suelen ser de cresta digitiformes (glándulas adrenales).
(a) Fotogralía al microscopio electrónico de transmisión de una sección longitudinal de una mitocondna de célula pancreática de rata (34.000x). (b) Representación tridimensional de una mitocondria.
http://www.youtube.com/watch?v=ornB9_UG65A&feature=related
--> Membrana externa
Es una bicapa lipídica exterior permeable a iones, metabolitos y muchos polipéptidos. Eso es debido a que contiene proteínas que forman poros, llamadas porinas o VDAC (de canal aniónico dependiente de voltaje), que permiten el paso de grandes moléculas de hasta 10.000 dalton y un diámetro aproximado de 20 Å. La membrana externa realiza relativamente pocas funciones enzimáticas o de transporte. Contiene entre un 60 y un 70% de proteínas.
--> Membrana interna
La membrana interna contiene más proteínas, carece de poros y es altamente selectiva; contiene muchos complejos enzimáticos y sistemas de transporte transmembrana, que están implicados en la translocación de moléculas. Esta membrana forma invaginaciones o pliegues llamadas crestas mitocondriales, que aumentan mucho la superficie para el asentamiento de dichas enzimas. En la mayoría de los eucariontes, las crestas forman tabiques aplanados perpendiculares al eje de la mitocondria, pero en algunos protistas tienen forma tubular o discoidal. En la composición de la membrana interna hay una gran abundancia de proteínas (un 80%), que son además exclusivas de este orgánulo:
--> Espacio intermembranoso
Entre ambas membranas queda delimitado un espacio intermembranoso está compuesto de un líquido similar al hialoplasma; tienen una alta concentración de protones como resultado del bombeo de los mismos por los complejos enzimáticos de la cadena respiratoria. En él se localizan diversos enzimas que intervienen en la transferencia del enlace de alta energía del ATP, como la adenilato quinasa o la creatina quinasa.
--> Matriz mitocondrial
La matriz mitocondrial o mitosol contiene menos moléculas que el citosol, aunque contiene iones, metabolitos a oxidar, ADN circular bicatenario muy parecido al de las bacterias, ribosomas tipo 70S similares a los de bacterias, llamados mitorribosomas, que realizan la síntesis de algunas proteínas mitocondriales, y contiene ARN mitocondrial; es decir, tienen los orgánulos que tendría una célula procariota de vida libre. En la matriz mitocondrial tienen lugar diversas rutas metabólicas clave para la vida, como el ciclo de Krebs y la beta-oxidación de los ácidos grasos; también se oxidan los aminoácidos y se localizan algunas reacciones de la síntesis de urea y grupos hemo.
* CURIOSIDADES:
· Relación de las mitocondrias con la teoría endosimbiótica:
Las mitocondrias son estructuras muy plásticas que se deforman, se dividen y fusionan. Normalmente se las representa en forma alargada. Su tamaño oscila entre 0,5 y 1 μm de diámetro y hasta 7 μ de longitud.Su número depende de las necesidades energéticas de la célula. Al conjunto de las mitocondrias de la célula se le denomina condrioma celular.
Las mitocondrias, orgánulos esféricos o alargados, aparecen repartidas por el citoplasma de las células eucarióticas . Una mitocondria está formada por una doble membrana de estructura similar a la de la membrana citoplasmática. La membrana mitocondrial externa es lisa, pero la interna se dispone formando una serie de pliegues llamadas crestas. El centro de la mitocondria se denomina matriz. Además poseen un espacio intermembranoso.Algunas enzimas implicadas en la respiración celular, incluyendo las que forman ATP, están localizadas sobre las crestas de la membrana interna mitocondrial y muchos de los pasos metabólicos relacionados con la respiración celular se realizan en la matriz.
Su número varía ampliamente según el tipo de organismo o tejido. A
lgunas células carecen de mitocondrias o poseen sólo una, mientras que otras pueden contener varios miles.Este orgánulo se compone de compartimentos que llevan a cabo funciones especializadas. Las proteínas mitocondriales varían dependiendo del tejido y de las especies: en humanos se han identificado 615 tipos de proteínas distintas en mitocondrias de músculo cardíaco;mientras que en ratas se han publicado 940 proteínas codificadas por distintos genes. Se piensa que el proteoma mitocondrial está sujeto a regulación dinámica.Aunque la mayor parte del ADN de la célula está en el núcleo celular, la mitocondria tiene su propio genoma, que muestra muchas semejanzas con los genomas bacterianos.Las mitocondrias tienen las crestas laminares o digitiformes.
La mayoría son de crestas laminares, mientras que las mitocondrias de células productoras de estrógenos suelen ser de cresta digitiformes (glándulas adrenales).
(a) Fotogralía al microscopio electrónico de transmisión de una sección longitudinal de una mitocondna de célula pancreática de rata (34.000x). (b) Representación tridimensional de una mitocondria.
http://www.youtube.com/watch?v=ornB9_UG65A&feature=related
--> Membrana externa
Es una bicapa lipídica exterior permeable a iones, metabolitos y muchos polipéptidos. Eso es debido a que contiene proteínas que forman poros, llamadas porinas o VDAC (de canal aniónico dependiente de voltaje), que permiten el paso de grandes moléculas de hasta 10.000 dalton y un diámetro aproximado de 20 Å. La membrana externa realiza relativamente pocas funciones enzimáticas o de transporte. Contiene entre un 60 y un 70% de proteínas.
--> Membrana interna
La membrana interna contiene más proteínas, carece de poros y es altamente selectiva; contiene muchos complejos enzimáticos y sistemas de transporte transmembrana, que están implicados en la translocación de moléculas. Esta membrana forma invaginaciones o pliegues llamadas crestas mitocondriales, que aumentan mucho la superficie para el asentamiento de dichas enzimas. En la mayoría de los eucariontes, las crestas forman tabiques aplanados perpendiculares al eje de la mitocondria, pero en algunos protistas tienen forma tubular o discoidal. En la composición de la membrana interna hay una gran abundancia de proteínas (un 80%), que son además exclusivas de este orgánulo:
--> Espacio intermembranoso
Entre ambas membranas queda delimitado un espacio intermembranoso está compuesto de un líquido similar al hialoplasma; tienen una alta concentración de protones como resultado del bombeo de los mismos por los complejos enzimáticos de la cadena respiratoria. En él se localizan diversos enzimas que intervienen en la transferencia del enlace de alta energía del ATP, como la adenilato quinasa o la creatina quinasa.
--> Matriz mitocondrial
La matriz mitocondrial o mitosol contiene menos moléculas que el citosol, aunque contiene iones, metabolitos a oxidar, ADN circular bicatenario muy parecido al de las bacterias, ribosomas tipo 70S similares a los de bacterias, llamados mitorribosomas, que realizan la síntesis de algunas proteínas mitocondriales, y contiene ARN mitocondrial; es decir, tienen los orgánulos que tendría una célula procariota de vida libre. En la matriz mitocondrial tienen lugar diversas rutas metabólicas clave para la vida, como el ciclo de Krebs y la beta-oxidación de los ácidos grasos; también se oxidan los aminoácidos y se localizan algunas reacciones de la síntesis de urea y grupos hemo.
* CURIOSIDADES:
· Relación de las mitocondrias con la teoría endosimbiótica:
Hace 2 billones de años hubo un desastre global que cambio las condiciones del planeta tierra por completo. La atmósfera contenía grandes cantidades de Dióxido de Carbono y pequeñas cantidades de Azufre, Nitrógeno, Carbono y Oxígeno. Después del desastre, los niveles de oxígeno en el ambiente empezaron a subir, probablemente por la acción de bacterias fotosintéticas del oceano. Al acumularse el oxígeno en tan grandes cantidades, mucha de la vida anaeróbica no logró protegerse o esconderse de la nueva atmósfera oxidativa por lo que se extinguieron. Otros organismos lograron protegerse de este oxígeno reduciendo las concentraciones al metabolizarlo y produciendo energía en forma de ATP.
Se piensa que se estableció una simbiosis entre una mitocondria y una célula huésped anaeróbica, a la cual le servía que la mitocondria eliminara el oxígeno del ambiente.
La enzima superoxido dismutasa es la máquina más rápida de nuestro cuerpo, se encarga de eliminar los radicales libres del oxigeno producidos por error en las mitocondrias. Es capaz de realizar hasta 7.000 millones de ciclos en un segundo, estando sólo limitada por la difusión de los sustratos. Al eliminar estos radicales libres impide que las células sean dañadas por el estrés oxidativo, el cual es, en parte, responsable del envejecimiento celular.
· Las mitocondrias se utilizan para buscar los ancestros de organismos que contienen células eucarióticas. Entre los mamíferos, las mitocondrias tienden a seguir una pauta de herencia materna.
Cuando una célula se divide, las mitocondrias se reproducen con independencia del núcleo. Las dos células hijas formadas después de la división reciben cada una la mitad de las mitocondrias. Cuando el espermatozoide fecunda al óvulo, sus mitocondrias quedan fuera del huevo. El cigoto fecundado hereda sólo las mitocondrias de la madre. Esta herencia materna crea un árbol familiar que no se ve afectado por la recombinación de genes que tiene lugar entre el padre y la madre.
Una comparación reciente de muestras de ADNm humano sugiere que la humanidad desciende de una mujer que vivió en África hace entre 140.000 y 290.000 años. Muestras genéticas tomadas de grupos étnicos africanos, asiáticos, australianos, europeos y de Nueva Guinea han revelado un número específico de tipos de ADNm. La comparación de estos tipos ha permitido a los científicos construir un árbol genealógico que sugiere que los distintos grupos empezaron probablemente a evolucionar por separado. En este árbol, el ADNm africano ocupa la rama más larga y antigua y de ella brotan los demás grupos étnicos. Probablemente había muchas otras mujeres vivas en la época de la llamada Eva mitocondrial, pero sus líneas de herencia materna se han extinguido. Esto ocurre habitualmente cuando una generación de una familia no produce ninguna hija.
· Según investigaciones recientes, unas pocas enfermedades de origen mitocondrial (heredadas por línea materna) son imputables a defectos del ADNm, entre ellas algunas patologías neuromusculares y ciertas formas de diabetes mellitas.
· El ADN mitocondrial puede dañarse con los radicales libres formados en la mitocondria; así, enfermedades degenerativas relacionadas con el envejecimiento, como la enfermedad de Parkinson, la enfermedad de Alzheimer y las cardioptías pueden tener relaciones con lesiones mitocondriales
miércoles, 17 de febrero de 2010
MITOSIS Y CITOCINESIS

En biología, la mitosis es un proceso de reparto equitativo del material hereditario (ADN) característico de las células eucarióticas. Normalmente concluye con la formación de dos núcleos separados (cariocinesis), seguido de la partición del citoplasma (citocinesis), para formar dos células hijas. La mitosis completa, que produce células genéticamente idénticas, es el fundamento del crecimiento, de la reparación tisular y de la reproducción asexual. No se debe confundir con la meiosis.
La mitosis es el tipo de división celular por el cual se conservan los orgánulos y la información genética contenida en sus cromosomas, que pasa de esta manera a las células hijas resultantes de la mitosis. La mitosis es igualmente un verdadero proceso de multiplicación celular que participa en el desarrollo, el crecimiento y la regeneración del organismo. Este proceso tiene lugar por medio de una serie de operaciones sucesivas que se desarrollan de una manera continua, y que para facilitar su estudio han sido separadas en varias etapas.
El resultado esencial de la mitosis es la continuidad de la información hereditaria de la célula madre en cada una de las dos células hijas. El genoma se compone de una determinada cantidad de genes organizados en cromosomas, hebras de ADN muy enrolladas que contienen la información genética vital para la célula y el organismo. Dado que cada célula debe contener completa la información genética propia de su especie, la célula madre debe hacer una copia de cada cromosoma antes de la mitosis, de forma que las dos células hijas reciban completa la información. Esto ocurre durante la fase S de la interfase, el período que alterna con la mitosis en el ciclo celular y en el que la célula entre otras cosas se prepara para dividirse.
Tras la duplicación del ADN, cada cromosoma consistirá en dos copias idénticas de la misma hebra de ADN, llamadas cromátidas hermanas, unidas entre sí por una región del cromosoma llamada centrómero. Cada cromátida hermana no se considera en esa situación un cromosoma en sí mismo, sino parte de un cromosoma que provisionalmente consta de dos cromátidas.
Se llama cariocinesis a la formación de los dos núcleos con que concluye habitualmente la mitosis. Es posible, y ocurre en ciertos casos, que el reparto mitótico se produzca sin cariocinesis (endomitosis) dando lugar a un núcleo con el material hereditario duplicado (doble número de cromosomas).
Cabe señalar que las células procariotas experimentan un proceso similar a la mitosis llamado fisión binaria. No se puede considerar que las células procariotas experimenten mitosis, dado que carecen de núcleo y únicamente tienen un cromosoma sin centrómero.
La mitosis es el tipo de división celular por el cual se conservan los orgánulos y la información genética contenida en sus cromosomas, que pasa de esta manera a las células hijas resultantes de la mitosis. La mitosis es igualmente un verdadero proceso de multiplicación celular que participa en el desarrollo, el crecimiento y la regeneración del organismo. Este proceso tiene lugar por medio de una serie de operaciones sucesivas que se desarrollan de una manera continua, y que para facilitar su estudio han sido separadas en varias etapas.
El resultado esencial de la mitosis es la continuidad de la información hereditaria de la célula madre en cada una de las dos células hijas. El genoma se compone de una determinada cantidad de genes organizados en cromosomas, hebras de ADN muy enrolladas que contienen la información genética vital para la célula y el organismo. Dado que cada célula debe contener completa la información genética propia de su especie, la célula madre debe hacer una copia de cada cromosoma antes de la mitosis, de forma que las dos células hijas reciban completa la información. Esto ocurre durante la fase S de la interfase, el período que alterna con la mitosis en el ciclo celular y en el que la célula entre otras cosas se prepara para dividirse.
Tras la duplicación del ADN, cada cromosoma consistirá en dos copias idénticas de la misma hebra de ADN, llamadas cromátidas hermanas, unidas entre sí por una región del cromosoma llamada centrómero. Cada cromátida hermana no se considera en esa situación un cromosoma en sí mismo, sino parte de un cromosoma que provisionalmente consta de dos cromátidas.
Se llama cariocinesis a la formación de los dos núcleos con que concluye habitualmente la mitosis. Es posible, y ocurre en ciertos casos, que el reparto mitótico se produzca sin cariocinesis (endomitosis) dando lugar a un núcleo con el material hereditario duplicado (doble número de cromosomas).
Cabe señalar que las células procariotas experimentan un proceso similar a la mitosis llamado fisión binaria. No se puede considerar que las células procariotas experimenten mitosis, dado que carecen de núcleo y únicamente tienen un cromosoma sin centrómero.
PROFASE

Los dos centros de origen de los microtúbulos son los centrosomas. La cromatina ha comenzado a condensarse y se observan las cromátidas. Los cinetocoros comienzan a formarse.
Es la fase más larga de la mitosis. Se produce en ella la condensación del material genético (ADN, que en interfase existe en forma de cromatina), para formar unas estructuras altamente organizadas, los cromosomas. Como el material genético se ha duplicado previamente durante la fase S, los cromosomas replicados están formados por dos cromátidas, unidas a través del centrómero por moléculas de cohesinas.
Uno de los hechos más tempranos de la profase en las células animales es duplicación del centrosoma; los dos centrosomas hijos (cada uno con dos centriolos) migran entonces hacia extremos opuestos de la célula. Los centrosomas actúan como centros organizadores de microtúbulos, controlando la formación de unas estructuras fibrosas, los microtúbulos, mediante la polimerización de tubulina soluble. De esta forma, el huso de una célula mitótica tiene dos polos que emanan microtúbulos.
En la profase tardía desaparece el nucléolo y se desorganiza la envoltura nuclear.
Uno de los hechos más tempranos de la profase en las células animales es duplicación del centrosoma; los dos centrosomas hijos (cada uno con dos centriolos) migran entonces hacia extremos opuestos de la célula. Los centrosomas actúan como centros organizadores de microtúbulos, controlando la formación de unas estructuras fibrosas, los microtúbulos, mediante la polimerización de tubulina soluble. De esta forma, el huso de una célula mitótica tiene dos polos que emanan microtúbulos.
En la profase tardía desaparece el nucléolo y se desorganiza la envoltura nuclear.
PROMETAFASE

La membrana nuclear se ha disuelto, y los microtúbulos invaden el espacio nuclear. Los microtúbulos pueden anclar cromosomas a través de los cinetocoros o interactuar con microtúbulos emanados por el polo opuesto.
La membrana nuclear se desensambla y los microtúbulos invaden el espacio nuclear. Esto se denomina mitosis abierta, y ocurre en una pequeña parte de los organismos multicelulares. Los hongos y algunos protistas, como las algas o las tricomonas, realizan una variación denominada mitosis cerrada, en la que el huso se forma dentro del núcleo o sus microtúbulos pueden penetrar a través de la membrana nuclear intacta.
Cada cromosoma ensambla dos cinetocoros hermanos sobre el centrómero, uno en cada cromátida. Un cinetocoro es una estructura proteica compleja a la que se anclan los microtúbulos. Aunque la estructura y la función del cinetocoro no se conoce completamente, contiene varios motores moleculares, entre otros componentes. Cuando un microtúbulo se ancla a un cinetocoro, los motores se activan, utilizando energía de la hidrólisis del ATP para "ascender" por el microtúbulo hacia el centrosoma de origen. Esta actividad motora, acoplada con la polimerización/despolimerización de los microtúbulos, proporcionan la fuerza de empuje necesaria para separar más adelante las dos cromátidas de los cromosomas.
Cuando el huso crece hasta una longitud suficiente, los microtúbulos asociados a cinetocoros empiezan a buscar cinetocoros a los que anclarse. Otros microtúbulos no se asocian a cinetocoros, sino a otros microtúbulos originados en el centrosoma opuesto para formar el huso mitótico. La prometafase se considera a veces como parte de la profase.
La membrana nuclear se desensambla y los microtúbulos invaden el espacio nuclear. Esto se denomina mitosis abierta, y ocurre en una pequeña parte de los organismos multicelulares. Los hongos y algunos protistas, como las algas o las tricomonas, realizan una variación denominada mitosis cerrada, en la que el huso se forma dentro del núcleo o sus microtúbulos pueden penetrar a través de la membrana nuclear intacta.
Cada cromosoma ensambla dos cinetocoros hermanos sobre el centrómero, uno en cada cromátida. Un cinetocoro es una estructura proteica compleja a la que se anclan los microtúbulos. Aunque la estructura y la función del cinetocoro no se conoce completamente, contiene varios motores moleculares, entre otros componentes. Cuando un microtúbulo se ancla a un cinetocoro, los motores se activan, utilizando energía de la hidrólisis del ATP para "ascender" por el microtúbulo hacia el centrosoma de origen. Esta actividad motora, acoplada con la polimerización/despolimerización de los microtúbulos, proporcionan la fuerza de empuje necesaria para separar más adelante las dos cromátidas de los cromosomas.
Cuando el huso crece hasta una longitud suficiente, los microtúbulos asociados a cinetocoros empiezan a buscar cinetocoros a los que anclarse. Otros microtúbulos no se asocian a cinetocoros, sino a otros microtúbulos originados en el centrosoma opuesto para formar el huso mitótico. La prometafase se considera a veces como parte de la profase.
METAFASE

A medida que los microtúbulos encuentran y se anclan a los cinetocoros durante la prometafase, los centrómeros de los cromosomas se congregan en la "placa metafásica" o "plano ecuatorial", una línea imaginaria que es equidistante de los dos centrosomas que se encuentran en los dos polos del huso. Este alineamiento equilibrado en la línea media del huso se debe a las fuerzas iguales y opuestas que se generan por los cinetocoros hermanos. Dado que una separación cromosómica correcta requiere que cada cinetocoro esté asociado a un conjunto de microtúbulos (que forman las fibras cinetocóricas), los cinetocoros que no están anclados generan una señal para evitar la progresión prematura hacia anafase antes de que todos los cromosomas estén correctamente anclados y alineados en la placa metafásica. Esta señal activa el checkpoint de mitosis.
ANAFASE

Cuando todos los cromosomas están correctamente anclados a los microtúbulos del huso y alineados en la placa metafásica, la célula procede a entrar en anafase.
Entonces tienen lugar dos sucesos. Primero, las proteínas que mantenían unidas ambas cromatidas hermanas (las cohesinas), son cortadas, lo que permite la separación de las cromátidas. Estas cromátidas hermanas, que ahora son cromosomas hermanos diferentes, son separados por los microtúbulos anclados a sus microtúbulos al desensamblarse, dirigiéndose hacia los centrosomas respectivos.
A continuación, los microtúbulos no asociados a cinetocoros se alargan, empujando a los centrosomas (y al conjunto de cromosomas que tienen asociados) hacia los extremos opuestos de la célula. Este movimento parece estar generado por el rápido ensamblaje de los microtúbulos.
Estos dos estadios se denominan a veces anafase temprana (A) y anafase tardía (B). La anafase temprana viene definida por la separación de cromátidas hermanas, mientras que la tardía por la elongación de los microtúbulos que produce la separación de los centrosomas. Al final de la anafase, la célula ha conseguido separar dos juegos idénticos de material genético en dos grupos definidos, cada uno alrededor de un centrosoma.
A continuación, los microtúbulos no asociados a cinetocoros se alargan, empujando a los centrosomas (y al conjunto de cromosomas que tienen asociados) hacia los extremos opuestos de la célula. Este movimento parece estar generado por el rápido ensamblaje de los microtúbulos.
Estos dos estadios se denominan a veces anafase temprana (A) y anafase tardía (B). La anafase temprana viene definida por la separación de cromátidas hermanas, mientras que la tardía por la elongación de los microtúbulos que produce la separación de los centrosomas. Al final de la anafase, la célula ha conseguido separar dos juegos idénticos de material genético en dos grupos definidos, cada uno alrededor de un centrosoma.
TELOFASE

La telofase es la reversión de los procesos que tuvieron lugar durante profase y prometafase. Durante la telofase, los microtúbulos no unidos a cinetocoros continúan alargándose, estirando aún más la célula. Los cromosomas hermanos se encuentran cada uno asociado a uno de los polos. La membrana nuclear se reforma alrededor de ambos grupos cromosómicos, utilizando fragmentos de la membrana nuclear de la célula original. Ambos juegos de cromosomas, ahora formando dos nuevos núcleos, se descondensan de nuevo en cromatina. La cariocinesis ha terminado, pero la división celular aún no está completa.
CITOCINÉSIS

La citocinesis es un proceso independiente, que se inicia simultáneamente a la telofase. Técnicamente no es parte de la mitosis, sino un proceso aparte, necesario para completar la división celular. En las células animales, se genera un surco de escisión (cleavage furrow) que contiene un anillo contráctil de actina en el lugar donde estuvo la placa metafásica, estrangulando el citoplasma y aislando así los dos nuevos núcleos en dos células hijas. Tanto en células animales como en plantas, la división celular está dirigida por vesículas

derivadas del aparato de Golgi, que se mueven a lo largo de los microtúbulos hasta la zona ecuatorial de la célula. En plantas esta estructura coalesce en una placa celular en el centro del fragmoplasto y se desarrolla generando una pared celular que separa los dos núcleos. El fragmoplasto es una estructura de microtúbulos típica de plantas superiores, mientras que algunas algas utilizan un vector de microtúbulos denominado ficoplasto durante la citocinesis. Al final del proceso, cada célula hija tiene una copia completa del genoma de la célula original. El final de la citocinesis marca el final de la fase M.
Consecuencias de la mitosis
Mediante el proceso mitótico, el material genético se divide en dos núcleos idénticos, con lo que las dos células hijas que resultan si se produce la división del citoplasma serán genéticamente idénticas. Por tanto, la mitosis es un proceso de división conservativo, ya que el material genético se mantiene de una generación celular a la siguiente. La mayor parte de la expresión génica se detiene durante la mitosis, pero mecanismos epigenéticos funcionan durante esta fase, para "recordar" los genes que estaban activos en mitosis y transmitirlos a las células hijas.
Errores en la mitosis
Aunque los errores en la mitosis son bastante poco frecuentes, este proceso puede fallar, especialmente durante las primeras divisiones celulares en el cigoto. Los errores mitóticos pueden ser especialmente peligrosos para el organismo, porque el descendiente futuro de la célula madre defectuosa mantendrá la misma anomalía.
Un cromosoma puede no separarse durante la anafase. Este fenómeno se denomina "no-disyunción". Si esto ocurre, una célula hija recibirá dos cromosomas hermanos y la otra se quedará sin ninguno. Esto da lugar a que una célula tenga tres cromosomas que codifiquen la misma cosa (dos hermanos y un homólogo), una condición conocida como trisomía, y la otra célula, que solamente tiene un cromosoma (el cromosoma homólogo), tendrá monosomía. Estas células se consideran aneuploides, y la aneuploidía puede causar inestabilidad genética, un hecho frecuente en cáncer.
La mitosis es un proceso traumático. La célula pasa por cambios drásticos en su estructura, algunos orgánulos se desintegran y se reconstruyen en cuestión de horas, y los microtúbulos tiran constantemente de los cromosomas. Por tanto, en ocasiones los cromosomas pueden dañarse. Un brazo del cromosoma se puede romper y perder un fragmento, causando deleción. El fragmento puede incorporarse incorrectamente a otro cromosoma no homólogo, causando translocación. Se puede integrar de nuevo al cromosoma original, pero en una orientación inversa, causando inversión. O se puede tratar erróneamente como un cromosoma separado, causando duplicación cromosómica.
Una parte de estos errores pueden detectarse por alguno de los puntos de control existentes a través del ciclo celular, lo cual produce una parada en la progresión celular, dando tiempo a los mecanismos reparadores a corregir el error. Si esto no ocurre, el efecto de estas anormalidades genéticas dependerá de la naturaleza específica del error. Puede variar de una anomalía imperceptible, a carcinogénesis o a la muerte del organismo.
Endomitosis
La endomitosis es una variante de la mitosis sin división nuclear o celular, lo que dá lugar a células con muchas copias del mismo cromosoma en el mismo núcleo. Este proceso también se denomina endoreduplicación, y las células resultantes endoploides. Un ejemplo de una célula que sufre endomitosis es el megacariocito.
Núcleo interfásico
http://www.youtube.com/watch?v=O6apHYhilLU
El núcleo puede considerarse como el orgánulo principal de la cálula eucariota, ya que en él reside el material genético y desde él se controla toda la actividad celular.
- Características del núcleo interfasico:
Los nucleos celulares tienen habitualmente forma esférica arriñonada o elipsoidal, aunque puede presentar otras menos frecuentes como la arrosariada o lilobulada.
Suele situarse en el centro de la célula, aunque hay numerosas excepciones como los núcleos laterales de las células adiposas.
Por regla general, el tamaño es proporcional al de la célula, aunque existe un límite. Su tamaño medio es de 8 a 10 umas, y normalmente ocupa un 10% o más del volumen celular. La relación existente entre el volumen nuclear y el citoplasmático se expresa mediante un coeficiente k, que viene definido por la fórmula:
Cuando k alcanza un cierto valor mínimo variable según el tipo de célula, esta entra en división. Es lo que ocurre cuando el citoplasma alcanza un cierto volumen: la célula se divide, como si el núcleo no pudiera controlar todo el volumen que le rodea.
Normalmente el núcleo tiene un núcleopero pueden tener más
- Composición química:
Los principales componentes encontrados han sido desoxirribonucleoproteinas, ribonucleoproteínas, enzimas, ARN solubles, nucleótidos de ADN y ARN, ATP, lípidos, aminoácidos, sales de magnesio, calcio, cinc y agua.
Las cantidades en las que aparecen estos compuestos pueden variar en función del tipo de célula. Así, en lo se refiere al ADN, por ejemplo, su proporción varia desde un 60% en los núcleos de los espermatozoides a un 1% en los ovocitos. Sin embarga, las cantidades son prácticamente la misma en cada uno de los tejidos de una misma especie.
- Actividad celular:
El núcleo interfásico desarrolla una intensa actividad metabólica que controla la vida de la célula. En él se produce tanto la actividad autosintética del ADN (replicación), como la heterosintetica, mediante la transcripción de la información genética al ARNm y la síntesis de los otros tipos de ARN y su ensamblamiento con las proteínas. Además, en el núcleo se lleva a cabo parte de la glucólisis.
Estas funciones, sin embargo, no podrían llevarse a cobo sin el citoplasma, del que recibe tanto las materias primas (sustratos y enzimas), como la energía, ya que el núcleo carece de una fuente autónoma de esta. La energía nuclear depende, pues, de los ATP, GTP, CTP y UTP citoplasmáticos.
La función de control del núcleo se pone de manifiesto en experiencias como la merotomia. Cuando una ameba se fracciona en dos, el fragmento del núcleo crece y sigue su vida pero el que no lo tiene muere, a no ser que se le trasplante el núcleo de otra ameba.
ENVOLTURA NUCLEAR:
La envoltura nuclear es una estructura compleja que se basa en una vesícula de retículo endoplasmático extendida alrededor del material hereditario nuclear (cromatina). Como tal vesícula, la envoltura aparece conformada por dos membranas: la membrana nuclear externa y la membrana nuclear interna. Por el lado de fuera queda el citoplasma y por el de dentro el contenido del núcleo. Por el lado del núcleo la membrana nuclear interna lleva adosada una estructura llamada lámina nuclear, la cual está formada por proteínas, como las llamadas laminas, a veces en forma de capa continua, a veces con la estructura de un panal. El hecho de que la envoltura sea una especialización del retículo endoplasmático se observa también en que suele aparecer recubierta de ribosomas (algo que es característico del retículo endoplasmático rugoso), los cuales fabrican precisamente proteínas que se incorporan a la composición de las membranas nucleares. Entre ambas membranas se encuentra una cavidad conocida como espacio cisternal o perinuclear, más ancho que cada membrana (las membranas miden de 7 a 8 nanómetros y el espacio entre ella de 10 a 20).

Principalmente delimita dos compartimentos funcionales dentro de la célula misma, el de transcripción ADN en ARN (dentro del nucleo) y el de traducción ARN en Proteína (en el citoplasma). La envoltura nuclear aparece atravesada d
 e manera regular por perforaciones, los poros nucleares. Estos poros no son simples orificios, sino estructuras complejas acompañadas de una armazón de proteínas (por ejemplo: nucleoporina), que facilitan a la vez que regulan los intercambios entre el núcleo y el citoplasma. Se llama complejo del poro a cada una de esas puertas de comunicación. Por ahí salen las moléculas de ARNm producidas por la transcripción, que deben ser leídas por los ribosomas del citoplasma. Por ahí salen también los complejos de ARNr y proteínas a partir de los cuales se ensamblan en el citoplasma los ribosomas. Por los poros entran al núcleo las proteínas, fabricadas en el citoplasma por los ribosomas, que cumplen su papel dentro del núcleo de la célula. Hay cerca de 2000 complejos de poro en la envoltura nuclear en la célula de un vertebrado, pero varía dependiendo del número de transcripciones de la célula.
e manera regular por perforaciones, los poros nucleares. Estos poros no son simples orificios, sino estructuras complejas acompañadas de una armazón de proteínas (por ejemplo: nucleoporina), que facilitan a la vez que regulan los intercambios entre el núcleo y el citoplasma. Se llama complejo del poro a cada una de esas puertas de comunicación. Por ahí salen las moléculas de ARNm producidas por la transcripción, que deben ser leídas por los ribosomas del citoplasma. Por ahí salen también los complejos de ARNr y proteínas a partir de los cuales se ensamblan en el citoplasma los ribosomas. Por los poros entran al núcleo las proteínas, fabricadas en el citoplasma por los ribosomas, que cumplen su papel dentro del núcleo de la célula. Hay cerca de 2000 complejos de poro en la envoltura nuclear en la célula de un vertebrado, pero varía dependiendo del número de transcripciones de la célula.Los poros están conectados entre si por la lámina nuclear. Esta lámina es una sustancia proteica que recubre la parte nucleoplasmática de la membrana interna, también llamada corteza nuclear o lámina fibrosa. Su espesor es de 50 a 80 nanómetros, y presenta el aspecto de una red fibrilar.

NUCLEOPLASMA:
El nucleoplasma o carioplasma es el medio interno del núcleo celular. Disolución coloidal que contiene gran cantidad de biomoléculas. En él se encuentran las fibras de ADN, que asociadas con proteínas denominadas histonas, forman hebras llamadas cromatinas y ARN conocidos como nucleólos. Contiene principalmente proteínas, sobre todo enzimas relacionados con el metabolismo de los ácidos nucléicos. También existen proteínas ácidas que no están unidas a ADN ni a ARN y que se denominan proteínas residuales. Además hay cofactores, moléculas precursoras, productos intermedios de la glucolisis, sodio, potasio, magnesio y calcio. También es el lugar dosde se produce la síntesis del ADN y ARN.
CROMATINA:
Es el conjunto de ADN, histonas y proteínas no histónicas que se encuentra en el núcleo de las células eucariotas y que constituye el cromosoma eucariótico. 
Las unidades básicas de la cromatina son los nucleosomas. Éstos se encuentran formados por aproximadamente 146 pares de bases de longitud (el número depende del organismo), asociados a un complejo específico de 8 histonas nucleosómicas (octámero de histonas). Cada partícula tiene una forma de disco, con un diámetro de 11 nm y contiene dos copias de cada una de las 4 histonas H3, H4, H2A y H2B. Este octámero forma un núcleo proteico alrededor del que se enrolla la hélice de ADN (da aproximadamente 1,8 vueltas). Entre cada una de las asociaciones de ADN e histonas existe un ADN libre llamado ADN "espaciador", de longitud variable entre 0 y 80 pares de nucleótidos que garantiza flexibilidad a la fibra de cromatina. Este tipo de organización, permite un primer paso de compactación del material genético, y da lugar a una estructura parecida a un "collar de cuentas".

Posteriormente, un segundo nivel de organización de orden superior lo constituye la "fibra de 30nm" compuestas por grupos de nucleosomas empaquetados uno sobre otros adoptando disposiciones regulares gracias a la acción de la histona H1.

Tipos de cromatina
Heterocromatina, es una forma inactiva condensada localizada sobre todo en la periferia del núcleo, que se tiñe fuertemente con las coloraciones.
En cuanto a la heterocromatina, la fibra que la constituye se encuentra más condensada y a menudo aparece formada por agregados. Su formación require numerosas proteínas adicionales, que incluyen las proteínas HP1 (Heterochromatin Protein 1 o proteína de la heterocromatina 1).
La heterocromatina puede ser de dos tipos diferentes :
La constitutiva, idéntica para todas las células del organismo y que carece de información genética, incluye a los telómeros y centrómeros del cromosoma que no expresan su ADN. Constitutiva contiene un tipo particular de ADN denominado ADN satélite, formado por gran número de secuencias cortas repetidas en tándem. Es estable y conserva sus propiedades heterocromáticas durante todas las etapas del desarrollo y en todos los tejidos. Es altamente polimórfica, probablemente debido a la inestabilidad del ADN satélite.
La facultativa, diferente en los distintos tipos celulares, contiene información sobre todos aquellos genes que no se expresan o que pueden expresarse en algún momento. Incluye al ADN satélite y al corpúsculo de Barr. Se caracteriza por la presencia de secuencias repetidas tipo LINE. Estas secuencias, dispersas a lo largo del genoma, podrían promover la propagación de una estructura de cromatina condensada. La heterocromatina facultativa es reversible, su estado heterocromático depende de la etapa del desarrollo y del tipo celular. La heterocromatina facultativa no es particularmente rica en ADN satélite, y por ello, no es polimórfica.
Propiedades de la heterocromatina: A pesar de las diferencias descritas anteriormente, la heterocromatina constitutiva y la heterocromatina facultativa tienen propiedades muy similares.
Eucromatina, está diseminada por el resto del núcleo. Representa la forma activa de la cromatina en la que se está transcribiendo el material genético de las moléculas de ADN a moléculas de ARNm, por lo que es aquí donde se encuentran la mayoría de los genes activos.
En la eucromatina inactiva, esta fibra se enrolla sobre sí misma gracias a las histonas H1 para formar el solenoide. La interacción con otras proteínas no histonas provoca mayores grados de organización.
Rol de la cromatina en la expresión génica:
La cromatina juega un rol regulatorio fundamental en la
. Por lo tanto, existe una variada cantidad de máquinas remodeladoras de la cromatina y modificadoras de histonas.
NUCLEOLO:
En biología celular, el nucleolo es una region del núcleo considerada como un orgánulo. La función principal del nucleolo es la producción y ensamblaje de los componentes ribosómicos. El nucleolo es aproximadamente esférico y está rodeado por una capa de cromatina condensada. El nucléolo, es la región heterocromatica más destacada del núcleo. No existe membrana que separe el nucleolo del nucleoplasma.
En biología celular, el nucleolo es una region del núcleo considerada como un orgánulo. La función principal del nucleolo es la producción y ensamblaje de los componentes ribosómicos. El nucleolo es aproximadamente esférico y está rodeado por una capa de cromatina condensada. El nucléolo, es la región heterocromatica más destacada del núcleo. No existe membrana que separe el nucleolo del nucleoplasma.
Los nucleolos están formados por proteínas y ADN ribosomal (ADNr). El ADNr es un componente fundamental ya que es utilizado como molde para la transcripción del ARN ribosómico, para incorporarlo a nuevos ribosomas. La mayor parte de las células tanto animales como vegetales, tienen uno o más nucleolos, aunque existen ciertos tipos celulares que no los tienen.

Morfológicamente, el nucléolo suele ser esférico pero puede adoptar formas muy irregulares. El nucléolo se divide en dos regiones:
Parte amorfa: se observa poco densa a los electrones está constituida por espacios intercomunicados entre sí y que quedan entre las partes más densas. Es equivalente al nucleoplasma.
Parte densa: forma el nucleolonema. Esta parte se observa densa a los electrones, pero existen diferentes regiones dependiendo de su grado de densidad:
Zona Central: es la región con menor densidad. Está formada por una red de fibrillas de 4-5 nanómetros de espesor. La forma es normalmente globular. En esta región se encuentre el ADN de los organizadores nucleolares y algunas proteínas y enzimas que intervienen en la transcripción.
Componentes Fibrilares Densos o Parte Fibrilar: es la región más densa. Son estructuras fibrilares de ribonucleoproteínas de un grosor de 8-10 nm y su tamaño refleja la cantidad de ARNr que se está produciendo.
Región granular: que se corresponden con las subunidades de ribosomas que se están formando. Los componentes granulares son pequeños gránulos con un diámetro de alrededor de 15 nm.
La función principal del nucléolo es la biogénesis de ribosomas desde su componentes de ADN para formar ARN prerribosomal. Está relacionado con la síntesis de proteínas. Además, investigaciones recientes, han descrito al nucléolo como el responsable del tráfico de pequeños segmentos de ARN. Además, interviene en la maduración y el transporte del ARN hasta su destino final en la célula. Aunque el nucléolo desaparezca en división, algunos estudios actuales aseguran que regula el ciclo celular.
Ciclo del nucléolo
El nucléolo no se observa a lo largo de todo el ciclo celular. Al igual que los cromosomas, sufre una serie de cambios según se encuentre en interfase o en división. En interfase no sufre cambios morfológicos significativos. Sin embargo en división se dan cambios que determinan el ciclo del nucléolo. En este ciclo hay tres etapas:
1. Desorganización profásica: el nucléolo disminuye de tamaño y se hace bastante irregular.
2. Transporte metafásico y anafásico: el nucléolo pierde su individualidad y sus componentes se incorporan a los cromosomas metafásicos.
3. Organización telofásica: en la primera mitad de la telofase, los cromosomas se descondensan y aparecen los cuerpos laminares y cuerpos prenucleolares. Estos cuerpos son estructuras esféricas con características citoquímicas y estructurales del núcleo interfásico. Los cuerpos prenucleolares aumentan de tamaño y empiezan a formar un nucléolo alrededor de laregión de los organizadores nucleolares. El número de nucléolos depende del número de organizadores nucleolares.
CURIOSIDADES:
CROMATINA: La Técnica de Azul de Anilina (A.A.) se usa para detectar defectos de la cromatina nuclear relacionadas con el contenido nucleo-proteico asociado con el ADN. En este test se evalúa el % de núcleos espermáticos maduros ricos en protaminas (cuya función principal es empaquetar el ADN en una cromatina muy condensada, proporcionando al núcleo espermático una extraordinaria estabilidad mecánica y química) que le dan al núcleo una gran estabilidad mecánica y química. Este test se usa para la eficacia de los espermatozoices en la reproducción sexual.
CROMATINA: La cromatina tiene un papel activo en la respuesta del organismo al estrés oxidativo ante la restricción calórica.La escasez de alimentos a la que se han tenido que enfrentar los seres vivos a lo largo de su historia evolutiva ha propiciado el desarrollo de diversas estrategias adaptativas de compensación que generan estrés oxidativo en los organismos, principalmente en las moléculas implicadas en el intercambio de información durante estos procesos. Este desequilibrio metabólico se ha asociado a diversas patologías como el cáncer, la diabetes y las enfermedades cardiovasculares. . Existen evidencias que apuntan a que las condiciones de escasez energética inducen una respuesta al estrés oxidativo, lo que implica un uso más eficiente de los recursos disponibles y la mejora del sistema de defensa frente a los efectos nocivos del estrés oxidativo.
Históricamente se había considerado que la cromatina era un actor pasivo en estos procesos, pero varios estudios recientes demuestran que la realidad es más compleja un papel muy activo en la prevención del daño celular producido por estrés oxidativo.
Históricamente se había considerado que la cromatina era un actor pasivo en estos procesos, pero varios estudios recientes demuestran que la realidad es más compleja un papel muy activo en la prevención del daño celular producido por estrés oxidativo.
CROMATINA: En la levadura Schizosaccharomyces pombe, el homólogo 6 de la proteína HP1 es absolutamente esencial para la cohesión eficiente de las cromátidas hermanas durante la división celular. Los experimentos en los cuales se ha realizado la deleción del ADN satellite muestran que una gran región de repeticiones de este tipo de ADN es indispensable para el funcionamiento correcto del centrómero.
NUCLEOLO: Después de 12 años de investigación, biólogos de la Facultad de Ciencias de la UNAM, rompieron un paradigma mundial, al demostrar que el microorganismo Giardia duodenalis sí tiene nucléolo, aunque es tan pequeño, que se ubica en la frontera entre lo que observan los microscópicos ópticos y los electrónicos.
Hasta ahora, se pensaba que el microorganismo estudiado era una especie de “eslabón perdido” entre el mundo de las bacterias o procariontes y procariontes.
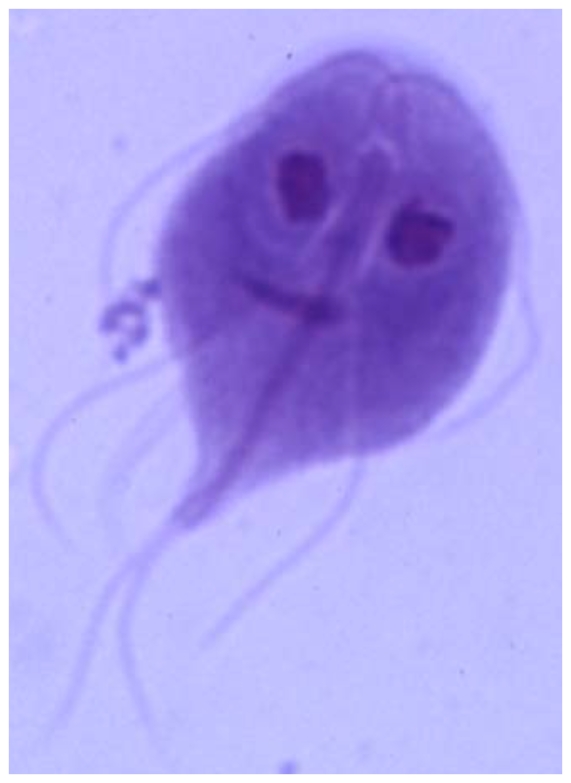
El nucléolo que se acostumbra estudiar mide dos, tres o cuatro micras, como ocurre en las células de los humanos o las plantas; pero en el caso de la Giardia fue difícil encontrarlo, pues mide 0.2 ó 0.3 micras, un tamaño inusual para un nucléolo.
martes, 16 de febrero de 2010
La Interfase
Interfase
Es el período de tiempo que transcurre entre dos mitosis, y que comprende los períodos G1, S, y G2. Durante la interfase se produce la duplicación de todos los componentes fundamentales de la célula, es decir DNA, RNA y proteínas; síntesis de lípidos, enzimas, membranas que se requieren para la división.

Es el período de tiempo que transcurre entre dos mitosis, y que comprende los períodos G1, S, y G2. Durante la interfase se produce la duplicación de todos los componentes fundamentales de la célula, es decir DNA, RNA y proteínas; síntesis de lípidos, enzimas, membranas que se requieren para la división.
El período G1, llamado primera fase de crecimiento, se inicia con una célula hija que proviene de la división de la célula madre. La célula aumenta de tamaño, se sintetiza nuevo material citoplásmico, sobre todo proteínas y RNA.
El período G0, la célula se encuentra en estado quiescente, cumpliendo con su función correspondiente, aunque en esta fase la célula no está preparándose para la división es donde llega la señal proteica.
El período S o de síntesis, en el que tiene lugar la duplicación del DNA. Cuando acaba este período, el núcleo contiene el doble de proteínas nucleares y de DNA que al principio. Hay síntesis de proteínas.

El período G2, DNA se sigue sintetizando RNA y proteínas; el final de este período queda marcado por la aparición de cambios en la estructura celular, que se hacen visibles con el microscopio y que nos indican el principio de la mitosis o división celular. Se realizan reparaciones en el DNA
El tiempo de cada fase es variable entre los organismos. En cada fase hay puntos de chequeo mediante proteínas que se fosforilizan o no.
La fase G1
Es el periodo del ciclo celular que abarca desde que termina la fase M hasta que comienza la fase S. Durante la fase G1 la célula comprueba las condiciones externas e internas y decide si continuar con el ciclo celular o no. En un organismo metazoo, el avance del ciclo celular está condicionado por señales externas, como adhesión, factores tróficos o mitógenos, entre otros, que emiten otras células del organismo. Las señales internas son aquellas que informan del estado de salud de la célula, como una correcta doctación de elementos celulares tras la división, una segregación correcta de los cromosomas, etcétera. Si todas estas señales son propicias la célula crecerá en tamaño y se preparará para entrar en la fase S.
Sin embargo, la mayoría de las células de un organismos pluricelular adulto no se dividen constantemente sino que detienen su ciclo celular en la fase G1, temporal o permanentemente. Detener el ciclo celular supone que la célula se va a diferenciar. Cuando la célula queda detenida en fase G1 en forma quiescente se dice que está en fase G0, un ejemplo son las neuronas. Desde los estados de quiescencia y de célula diferenciada se puede volver a retomar el ciclo en algunos tipos celulares. Por tanto tenemos cuatro decisiones posibles que se toman en la fase G1 y todas ellas dependen de complejos moleculares o puntos de control que la célula debe ir sorteando para llegar a la fase S. Cuando uno de ellos no se pasa se dice que la célula ha tomado una decisión, pero si no se detiene en ninguno se dividirá, siendo éste el camino por defecto.
Las moléculas que están en la base de los puntos de control, y por tanto de la progresión del ciclo celular, son las quinasas dependientes de ciclinas o CdKs (Cyclin-dependent kinases ). Estas enzimas, se han encontrado 9 diferentes en las células eucariotas, necesitan estar unidas a unas proteínas denominadas ciclinas y además ser activadas por fosforilación. Una vez activadas son las responsables de fosforilar numerosos sustratos, entre los que se encuentran los inhibidores del avance del ciclo celular, permitiendo así que el ciclo progrese. Las ciclinas son moléculas que se sintetizan de forma periódica durante el ciclo celular y se han encontrado hasta 16 ciclinas diferentes en las células eucariotas, siendo las más importantes para el avance del ciclo celular las A, B, D y E. Las ciclinas D y E son importantes para el avance de la fase G1. Los complejos CdK/ciclina D y Cdk/ciclina E actúan fosforilando al factor de transcripción Rb (retinoblastoma), que forma parte del último punto de control de la fase G1. Sin embargo, en levaduras sólo se require la actuación de
A este último punto de control en el que se fosforila a Rb se le denomina punto de restricción, porque si se pasa se entra irremediablemente en la fase S. Es importante porque se decide si la célula se dividirá o no. Los elementos centrales de este punto de restricción son
Pero, como dijimos, la mayoría de las células de un organismo adulto no están en permanente proliferación. Ello es debido a que existen inhibidores de las Cdk/ciclinas de la fase G1. Hay diversos tipos de inhibidores y uno de ellos es el p53, un factor de transcripción que está dañado en numerosos tipos de cánceres. Cuando hay daño del DNA celular, estrés celular, cambios de pH u otras alteraciones celulares, aumenta su concentración y provoca la activación del gen p21, el cual a su vez impide la fosforilación de Rb, y por tanto la célula no comienza la fase S.
La fase S
Comienza cuando se ha pasado el punto de restricción de la fase G1. Se producen dos sucesos importantes: replicación del ADN y duplicación de los centrosomas en las células animales.
Replicación del ADN
El ADN está formado por dos cadenas bases nucleotídicas. Ambas cadenas están unidas por puentes de hidrógeno que se establecen entre bases complementarias (adenina-timina, citosina-guanina), formando una doble hélice. Las dos cadenas se disponen de forma antiparalela entre sí. Esto quiere decir que el extremo 3' de una cadena está al lado del 5' de la otra cadena. Es decir, un extremo de la doble cadena posee un extremo 3' de una cadena y un extremo 5' de la otra. Para la duplicación del ADN hay que separar las dos cadenas rompiendo los puentes de hidrógeno y copiarlas simultáneamente.
El ADN de una célula eucariota no se copia empezando por un solo punto, esto llevaría demasiado tiempo, sino en múltiples sitios a la vez denominados orígenes de replicación. La célula dispone de los mecanismos necesarios para evitar que un origen de replicación se active más de una vez. Si no fuese así se producen más de una copia, lo que podría ser letal. Se consigue por un mecanimo en dos pasos. En el primero se organiza la maquinaria molecular necesaria para iniciar el proceso de copia y en segundo lugar se recibe una "licencia" para comenzar la replicación.
Para que se inicie la replicación se separan, no se rompen, las dos cadenas del ADN por una helicasa. A las cadenas libres se une una enzima denominada primasa que sintetizarán un pequeño fragmento de ARN de unos 10 nucleótidos complementarios a una secuencia de la cadena de ADN, uno distinto en cada cadena. A estas pequeñas secuencias de ARN se les denomina cebadores o "primers" Entonces se reclutan las polimerasas δ y ε, las cuales añadirán al extremo 3' desoxinucleótidos complementarios en la dirección del extremo 5' de la cadena copiada. Por tanto, formarán una cadena de nueva síntesis complementaria a cada una de las existentes previamente. Por eso se dice que la replicación es semiconservativa , una cadena nueva sobre una vieja. Un paso adicional es la eliminación del cebador de ribonucleótidos y su sustitución por desoxirribonucleótidos, llevado a cabo por ARNasas. El hueco se copiará por las DNA polimerasas que vienen copiando desde un origen de replicación situado más atrás en la cadena.
La apertura inicial de la doble cadena de ADN supone la creación de una horquilla de replicación. A partir de ella se copiarán las cadenas en las dos direcciones. Sin embargo, las ADN polimerasas añaden nucleódios exclusivamente en dirección 5' —» 3' (5' a 3' de la cadena copiada). Ello supone que la copia en la dirección 3' a 5' necesita de un proceso ligeramente más complicado. Así, en la zona de apertura de la doble hélice se irán añadiendo cebadores espaciados y serán los espacios entre estos cebadores los que llenarán las ADN polimerasas con nucleótidos complementarios pero siempre en dirección 3'. Esto supone que hay un proceso continúo de creación de cebadores, copia de ADN, eliminación de los cebadores más antiguos, copia del espacio dejado por ellos por las ADN polimerasas y sellado de los segmentos de ADN con las ezimas denominadas ligasas. A estos fragmentos de ADN, que se sintetizan periódicamente y son ligados entre sí para formar una cadena continua, se les denomina fragmentos de Okazaky.
Es importante tener en cuenta que no todo el ADN se está replicando a la vez. Se estima que en cualquier momento de la fase S se está copiando entre un 10 y un 15 % del ADN total. Si se detectan roturas del ADN, mediante los sistemas de control, la copia del resto del ADN se detiene. Otros eventos están ligados a la replicación del ADN como la síntesis de histonas, que debe también duplicar su número, y la duplicación de los centrosomas en las células animales, necesarios para organización del huso mitótico.
Fase G2
La fase S del ciclo celular da paso a la fase G2, la cual termina con la entrada en la fase M o mitosis. En la fase G2 se acumulan aquellas moléculas cuyas actividades serán necesarias durante la fase M. Tradicionalmente se ha considerado como un estado de tránsito entre las fases S y M. Durante esta etapa, sin embargo, se comprueba si ha habido errores durante la replicación del ADN y si se ha producido su duplicación completa. Si éstos defectos son detectados la célula no entrará en fase M y el ciclo celular se detendrá hasta que los daños sean reparados o el ADN sea completamente copiado. Se puede entender que estos mecanismos son críticos para la célula puesto que los errores no detectados pasarán irremediablemente a las células hijas. Durante la fase G2 la células también aumentarán en tamaño y los centrosomas, duplicados durante la fase S, se dirigirán a lugares opuestos de la célula para formar posteriormente el huso mitótico.
El límite entre las fases G2 y M no está totalmente claro y algunos autores consideran este cambio en la mitad de la profase mitótica. De cualquier manera, el fin de la fase G2 está mediado por la quinasa dependiente de ciclina (CdK) tipo 1 y por la ciclina B1. La ciclina B1 se sintetiza durante la fase S tardía. Es este complejo, más otras proteínas quinasas y fosfatasas, el que determina si la célula entrará en la fase M, es decir, es un punto de control.
Curiosidades
El cáncer es causado por anormalidades en el material genético de las células. Estas anormalidades pueden ser provocadas por agentes carcinógenos, de productos químicos o de agentes infecciosos. Otras anormalidades genéticas cancerígenas son adquiridas durante la replicación normal del ADN, al no corregirse los errores que se producen durante la misma, o bien son heredadas y, por consiguiente, se presentan en todas las células desde el nacimiento. Nuevos aspectos de la genética del cáncer, como la metilación del ADN, que es la adición de un grupo metilo (-CH3) a una molécula, consiste en la transferencia de grupos metilos a algunas de las bases citosinas (C) del ADN situadas previa y contiguamente a una guanina (G). Puesto que la metilación es fundamental en la regulación del silenciamiento de los genes, puede provocar alteraciones en la transcripción genética sin necesidad de que se produzca una alteración en la secuencia del ADN, siendo uno de los mecanismos responsables de la plasticidad fenotípica. También pueden ser metilados las proteínas, regulándose así también su función. En este proceso intervienen las enzimas ADN-metiltransferasas. Los microARNs, están siendo estudiados como importantes factores a tener en cuenta por su implicación. Hay muchos métodos para intentar acabar con todos los tipos de cáncer, pero hay algunos estudios nuevos relacionados con la genética como el estudio del ARN de la saliva, que podria acabar con el cancer oral. Los investigadores han descubierto al menos 50 microARNs en la saliva humana que podrían ayudar a la detección del cáncer oral.
pueden ser provocadas por agentes carcinógenos, de productos químicos o de agentes infecciosos. Otras anormalidades genéticas cancerígenas son adquiridas durante la replicación normal del ADN, al no corregirse los errores que se producen durante la misma, o bien son heredadas y, por consiguiente, se presentan en todas las células desde el nacimiento. Nuevos aspectos de la genética del cáncer, como la metilación del ADN, que es la adición de un grupo metilo (-CH3) a una molécula, consiste en la transferencia de grupos metilos a algunas de las bases citosinas (C) del ADN situadas previa y contiguamente a una guanina (G). Puesto que la metilación es fundamental en la regulación del silenciamiento de los genes, puede provocar alteraciones en la transcripción genética sin necesidad de que se produzca una alteración en la secuencia del ADN, siendo uno de los mecanismos responsables de la plasticidad fenotípica. También pueden ser metilados las proteínas, regulándose así también su función. En este proceso intervienen las enzimas ADN-metiltransferasas. Los microARNs, están siendo estudiados como importantes factores a tener en cuenta por su implicación. Hay muchos métodos para intentar acabar con todos los tipos de cáncer, pero hay algunos estudios nuevos relacionados con la genética como el estudio del ARN de la saliva, que podria acabar con el cancer oral. Los investigadores han descubierto al menos 50 microARNs en la saliva humana que podrían ayudar a la detección del cáncer oral.
Las anormalidades genéticas encontradas en las células cancerosas pueden ser de tipo mutación puntual, translocación, amplificación, deleción, y ganancia/pérdida de todo un cromosoma. Existen genes que son más susceptibles a sufrir mutaciones que desencadenen cáncer. Esos genes, cuando están en su estado normal, se llaman protooncogenes, y cuando están mutados se llaman oncogenes. Lo que esos genes codifican suelen ser receptores de factores de crecimiento, de manera que la mutación genética hace que los receptores producidos estén permanentemente activados, o bien codifican los factores de crecimiento en sí, y la mutación puede hacer que se produzcan factores de crecimiento en exceso y sin control.
Videos:
http://www.youtube.com/watch?v=_LRxFc21oXk&feature=related
http://www.youtube.com/watch?v=rgLJrvoX_qo
http://www.youtube.com/watch?v=unTshYSctAQ&NR=1
http://www.youtube.com/watch?v=ZV_o0jphro8&feature=related
Suscribirse a:
Comentarios (Atom)